Plant-pollinator systems present an ideal model for testing fundamental questions in biology. My research centers on how these systems can be leveraged to test how biological entities respond to environmental change. A prevailing theme is how reproduction – both whether reproduction occurs and, if so, via what reproductive strategy – depends on the environment. Bridging multiple scales of biological organization, my research helps elucidate how individual responses to the environment can affect population processes.
Pollen transfer as a bridge between ecology and evolution
Plant-pollinator systems exhibit a unique type of ecological interaction: plant reproduction depends on pollinator-mediated movement of gametes, while pollinator foraging depends on plants honestly signaling the availability of nutritional rewards. In turn, a system results where both partners affect the fitness and evolution of the other. Variation in floral phenotype affects the reliability that pollinators transfer intraspecific pollen between flowers, with unreliable pollen transfer driving pollen limitation and hybridization. My work explores this interplay between plants and pollinators: How does environmental variation affect the intra- and interspecific transfer of pollen, and what are the evolutionary consequences when pollen transfer is unreliable?
Pollen transfer depends on the foraging behavior of individual pollinators and floral community composition. My work has found that pollinators increase floral constancy (i.e., the tendency to make consecutive visits to the same floral species, thus increasing the likelihood of intraspecific pollen transfer) in environments with a higher number of co-flowering species (Fig. 1a) (Austin et al. 2019). Notably, my research has also revealed that flowering phenology shifts driven by climate change can increase co-flowering richness by protracting the duration of population-level flowering seasons (Fig. 1b) (Austin et al. in preparation). In my current work, I am leading a collaborative project that integrates archived specimen and time series field data with palynological techniques to reveal how such increased co-flowering richness has affected patterns of intra- and interspecific pollen transfer. Coupled with hand pollination experiments that assess the fitness effects of intra- and interspecific pollination, this work is revealing how pollen transfer mediates community composition and individual fitness in changing environments.
Pollen transfer depends on the foraging behavior of individual pollinators and floral community composition. My work has found that pollinators increase floral constancy (i.e., the tendency to make consecutive visits to the same floral species, thus increasing the likelihood of intraspecific pollen transfer) in environments with a higher number of co-flowering species (Fig. 1a) (Austin et al. 2019). Notably, my research has also revealed that flowering phenology shifts driven by climate change can increase co-flowering richness by protracting the duration of population-level flowering seasons (Fig. 1b) (Austin et al. in preparation). In my current work, I am leading a collaborative project that integrates archived specimen and time series field data with palynological techniques to reveal how such increased co-flowering richness has affected patterns of intra- and interspecific pollen transfer. Coupled with hand pollination experiments that assess the fitness effects of intra- and interspecific pollination, this work is revealing how pollen transfer mediates community composition and individual fitness in changing environments.
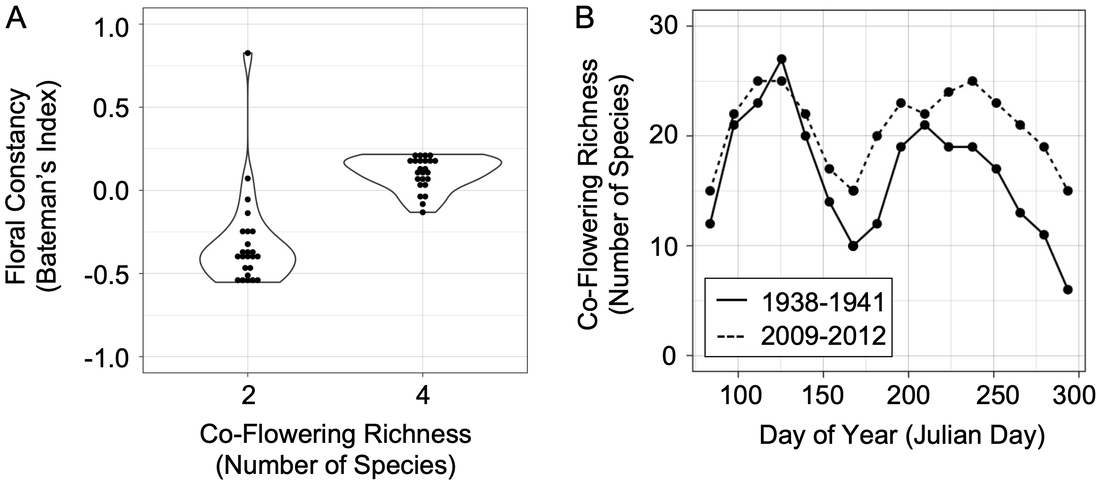
Figure 1. Co-flowering richness (A) increases bumble bee floral constancy (i.e., the tendency for a pollinator to make consecutive visits to the same floral species; P < 0.001) (Austin et al. 2019) and (B) has increased across the 20th century at Shaw Nature Reserve (Gray Summit, Missouri, USA; P < 0.001) (Austin et al. in preparation). Bateman’s index runs from -1 to 1, with higher values representing greater constancy. Panel A adapted from Austin et al. (2019).
Effects of climate change on mixed mating systems
In species with mixed mating systems (i.e., species that are nether exclusively outcrossing nor exclusively selfing), selfing is typically employed as a bet-hedging strategy in resource limited environments that are unsuitable for outcrossing. Accordingly, as climate change alters resource availability, species with mixed mating systems might respond with altered rates of outcrossing. My research has revealed that the common blue violet (Viola sororia; Violaceae) – a mixed mating species common across eastern North America – has responded to greater water availability in the North American Midwest by increasing investment in outcrossing (Austin et al. 2022).
I am developing V. sororia as a model for studying how climate change affects reproductive strategy investment in species with mixed mating systems. Viola sororia is a common species where investment in outcrossing versus selfing can be quantified by the production of morphologically distinct flowers (Fig. 2a,b); facultatively outcrossing chasmogamous (CH) flowers are produced under high precipitation and low temperature, while obligately selfing cleistogamous (CL) flowers are produced when precipitation is low and temperature is high (Fig. 2c). My work is demonstrating V. sororia as an ideal model for studying how species with mixed mating systems respond to climate change due to (1) its commonness across eastern North America, (2) the ease with which facultatively outcrossing and obligately selfing flowers can be identified, and (3) its amenability to study using a variety of field, lab, and herbarium techniques.
I am developing V. sororia as a model for studying how climate change affects reproductive strategy investment in species with mixed mating systems. Viola sororia is a common species where investment in outcrossing versus selfing can be quantified by the production of morphologically distinct flowers (Fig. 2a,b); facultatively outcrossing chasmogamous (CH) flowers are produced under high precipitation and low temperature, while obligately selfing cleistogamous (CL) flowers are produced when precipitation is low and temperature is high (Fig. 2c). My work is demonstrating V. sororia as an ideal model for studying how species with mixed mating systems respond to climate change due to (1) its commonness across eastern North America, (2) the ease with which facultatively outcrossing and obligately selfing flowers can be identified, and (3) its amenability to study using a variety of field, lab, and herbarium techniques.
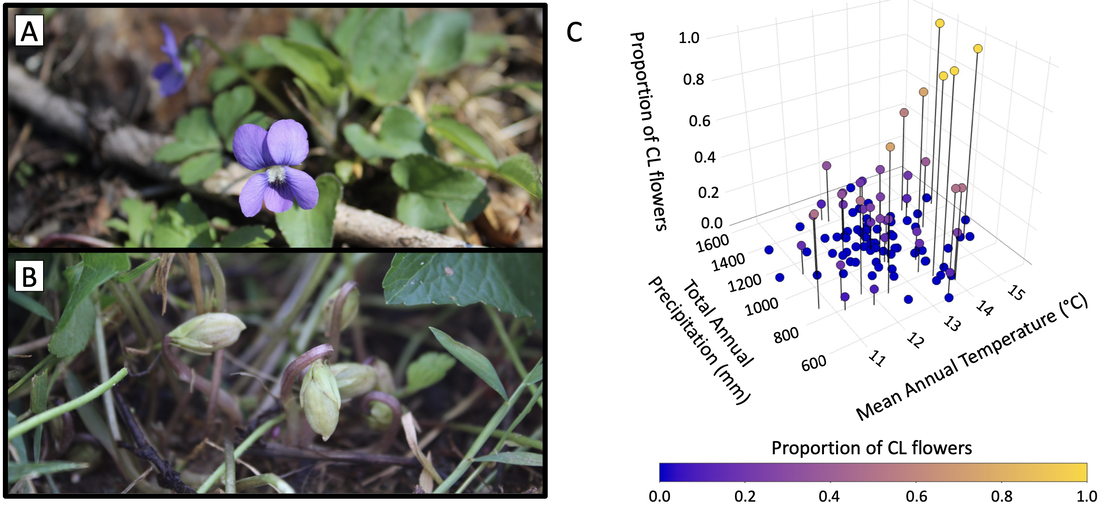
Figure 2. Mixed mating system of the common blue violet (Viola sororia; Violaceae). Viola sororia produces two types of morphologically distinct flowers: (A) chasmogamous (CH) flowers that facultatively outcross and (B) cleistogamous (CL) flowers that obligately self-fertilize. (C) CL flowers are produced at higher frequencies relative to CH flowers in environments with low total annual precipitation (P < 0.01) and high mean annual temperature (P < 0.05). Figure adapted from Austin et al. (2022).
The role of intraspecific variation in shaping population processes
Intraspecific trait variation is arguably one of the most important phenomena in biology. Trait variation is the raw material for adaptation by natural selection, enables evolutionary change in response to environmental changes, and trait plasticity can buffer populations from decline when confronted with environmental stochasticity. Yet biologists often overlook the role of intraspecific variation in shaping population processes and instead default to describing populations with mean trait values. My work is revealing the importance of intraspecific variation to understanding population-level processes. Bumble bees (Bombus spp.; Apidae) are the predominant model I have used for this work, which are an ideal system due to their high level of intraspecific trait variation, yet marked interspecific divergence. My work has demonstrated that bumble bee species with higher levels of intraspecific body size variation are less susceptible to population declines (Fig. 3) (Austin & Dunlap 2019). This is likely due to adaptive plasticity of body size across human modified environments (Austin et al. 2022) and a potential adaptive role of body size variation within colonies (Austin & Dunlap 2019). My work has also revealed that for species where cognition is developmentally plastic, resource phenology can drive seasonal changes to population-level cognition (Austin & Dunlap 2023). In my current work, I am exploring how intraspecific variation in flowering time and outcrossing rates affect intra- and interspecific gene flow, genetic diversity, and inbreeding depression.
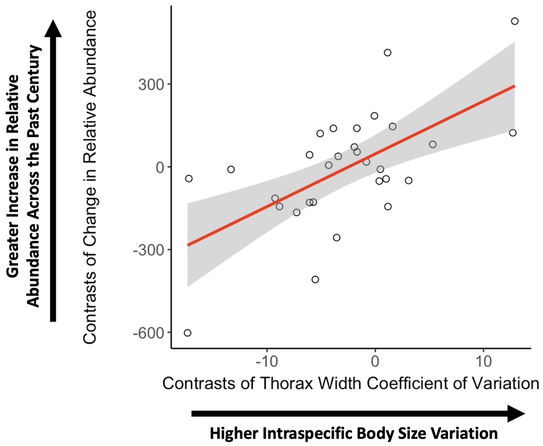
Figure 3. Intraspecific body size variation is positively correlated with changes to relative abundance across the past century for North American bumble bees (Bombus spp.; Apidae) (P < 0.001). Correlation depicts the phylogenetically controlled relationship between these variables. Gray area is the 95% confidence interval. Figure adapted from Austin & Dunlap (2019).
Figure 3. Intraspecific body size variation is positively correlated with changes to relative abundance across the past century for North American bumble bees (Bombus spp.; Apidae) (P < 0.001). Correlation depicts the phylogenetically controlled relationship between these variables. Gray area is the 95% confidence interval. Figure adapted from Austin & Dunlap (2019).
